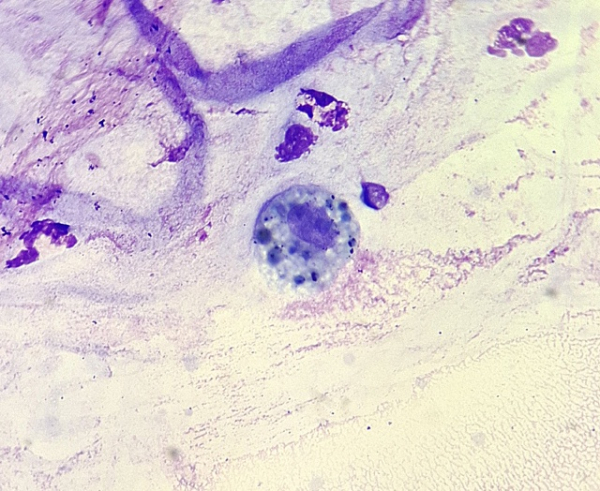

Макрофаги — это клетки, способные к активному захвату и перевариванию бактерий, остатков погибших клеток и других чужеродных или токсичных для организма частиц.
Устаревшие, вышедшие из употребления синонимы: гистиоцит-макрофаг, гистофагоцит, макрофагоцит, мегалофаг-пожиратель.
На протяжении нескольких веков макрофаги были в центре внимания клеточных биологов, патофизиологов, иммунологов и микробиологов. Макрофаги впервые были описаны как клетки, способные к поглощению инородных частиц или других клеток. Традиционно открытие макрофагов приписывается И.И. Мечникову, как и первое описание фагоцитоза. Однако, несмотря на широту распространения этого мнения, ряд авторов заявляют о несогласии с этим. Явление фагоцитоза впервые было описано немецким ученым Johann August Ephraim Goeze в 1777 году, чьи наблюдения подтвердил двумя годами позже Friederich Wilhelm von Gleichen-Russworm. Для клеток, способных к фагоцитозу, австрийский исследователь Carl Claus ввёл термин Fresszellen (поедающие клетки), который И.И. Мечников заменил термином «фагоциты». Макрофаги в нашем современном понимании, вероятно, впервые были описаны Giulio Bizzozero в 1871-1873 годах как клетки, участвующие в поглощении и разрушении эритроцитов, а также как клетки лимфатических узлов, поглощающие инородные частицы, попавшие в лимфу.
Таким образом, при изучении макрофагов с самого начала их описания преобладал функциональный подход, основанный на их способности к фагоцитозу. С использованием этого подхода были описаны многие резидентные (тканевые) макрофаги, в том числе клетки Купфера, альвеолярные макрофаги, макрофаги лимфатических узлов и др.
В настоящее время функциональный подход в обнаружении и исследовании различных популяций макрофагов сохранился, однако претерпел значительные изменения с учётом появления современных методов исследования. Фагоцитарная активность по-прежнему рассматривается как одно из ключевых свойств макрофагов, поэтому при изучении тех или иных популяций по-прежнему оценивают их фагоцитарную активность относительно латексных частиц, E. сoli и других бактерий.
В целом для макрофагов характерны несколько функций, касающихся воспаления и репарации:
фагоцитоз патогенов, дефектных и погибших клеток;
антиген-презентация;
продукция цитокинов;
участие в развитии воспалительных (сахарный диабет, рак, атеросклероз и др.) и пролиферативных заболеваний.
Анализируя участие макрофагов в воспалительных и репаративных реакциях, стало ясно, что данные клетки могут как стимулировать воспаление, так и приводить к его разрешению.
Исходя из этого в начале 2000-х годов в попытках классифицировать механизмы активации, формирования фенотипа, а также функциональные типы макрофагов Mills и соавт. по аналогии с Th1/Th2-иммунным ответом сформулировали М1/М2-парадигму активации макрофагов. В соответствии с М1/М2-дихотомией макрофаги разделены на М1 (провоспалительный) и М2 (противовоспалительный) типы.
Провоспалительные свойства макрофагов активируются при распознавании патоген-ассоциированных паттернов, что приводит к активации синтеза и выделению провоспалительных цитокинов, включая TNF?, IL-1?, IL-6, повышению экспрессии CD86 (является лигандом костимулятора Т-лимфоцитов CD28 или их ингибитора CTLA-4). Развитие воспалительной реакции способствует рекрутированию большего количества макрофагов и лейкоцитов. Однако развивающаяся воспалительная реакция оказывает негативное влияние на репаративные процессы в области повреждения, в связи с этим необходима альтернативная активация макрофагов, приобретение противовоспалительного фенотипа.
Приобретение макрофагами противовоспалительного фенотипа происходит в результате активации сигнальных путей под влиянием цитокинов IL-4, IL-13, IL-10 и IL-33. Макрофаги с противовоспалительным фенотипом характеризуются низкой продукцией IL-12, высокой продукцией IL-10, TGF?, повышением экспрессии рецепторов CD163 (скавенджер-рецептор комплексов гемоглобин-гаптоглобин), CD206 (рецептор маннозы, при воспалении связывающий и удаляющий из кровотока и межклеточной жидкости гидролазы, активатор тканевого плазминогена, миелопероксидазу); а также способностью к фагоцитозу патогенных организмов, погибших клеток и продуктов распада; участием в восстановлении и ремоделировании ткани; способностью стимулировать ангиогенез.
В 1970-х годах была сформулирована гипотеза о системе мононуклеарных фагоцитов, в соответствии с которой макрофаги представляют собой конечную стадию дифференцировки моноцитов крови, которые, в свою очередь, происходят из мультипотентных стволовых клеток крови в костном мозге. Однако исследования, проведённые в 2008-2013 годах, показали, что макрофаги тканей взрослых мышей представлены двумя популяциями, которые различаются по своему происхождению, механизму поддержания численности и функциям. Первая популяция это тканевые, или резидентные макрофаги. Они происходят из эритромиелоидных предшественников (не имеющих отношения к стволовым клеткам крови) желточного мешка и эмбриональной печени и заселяют ткани на различных этапах эмбриогенеза. Резидентные макрофаги приобретают тканеспецифичные характеристики и поддерживают свою численность за счёт пролиферации in situ без какого-либо участия моноцитов. К долгоживущим тканевым макрофагам относят купферовские клетки печени, микроглию центральной нервной системы, альвеолярные макрофаги лёгких, перитонеальные макрофаги брюшной полости, клетки Лангерганса кожи, макрофаги красной пульпы селезёнки.
Вторая популяция представлена относительно короткоживущими макрофагами моноцитарного (костномозгового) происхождения. Относительное содержание таких клеток в ткани зависит от её типа и возраста организма. Так макрофаги костномозгового происхождения составляют менее 5% всех макрофагов головного мозга, печени и эпидермиса, небольшую долю макрофагов лёгких, сердца и селезёнки (однако эта доля увеличивается с возрастом организма) и большую часть макрофагов собственной пластинки слизистой оболочки кишечника. Количество макрофагов моноцитарного происхождения резко увеличивается при воспалении и нормализуется по его окончании.
Таким образом, моноцит как специфический этап в развитии может быть обнаружен в большинстве макрофагальных линий млекопитающих (в зависимости от того, какую популяцию макрофагов мы рассматриваем, это будут либо фетальные моноциты, либо моноциты костномозгового происхождения). Долгое время моноциты рассматривались как недостаточно дифференцированные клетки. В настоящее время такой взгляд практически преодолён в научной литературе. Моноцит рассматривается как высокоспециализированная клетка с конкретными свойствами, которые могут различаться в зависимости от того, какую субпопуляцию мы рассматриваем. Нет сомнения, что одним из главных свойств моноцитов является их способность к миграции. Именно данная функция определяет профиль экспрессии генов и перечень поверхностных маркеров моноцитов, делая их высокочувствительными клетками к изменяющимся условиям микроокружения.
Источник: vk.com
Источник: ai-news.ru
 2")